
Protein folding MSM: villin headpiece#
You will learn: how to take a set of unbiased MD trajectories of a small protein and build a Markov-state model that gives you the folded / unfolded macrostates, the folding timescale, and the folding rate.
Prerequisites:
HTMD installed.
wgetavailable on$PATH(or substitute your favourite downloader).Patience for a one-time ~1 GB dataset download.
The system#
The villin headpiece is a 35-residue protein with a single hydrophobic core - one of the smallest naturally-occurring autonomously folding domains. We use a set of unbiased explicit-solvent trajectories run at 360 K (above the melting point, so both folded and unfolded states are populated). Total aggregate ≈ 107 µs across 2137 short trajectories.
The flow#
The MSM pipeline you’ll follow:
Download + index the filtered trajectories with
simlist().Project the geometry onto a low-dimensional distance space with
MetricSelfDistance.Reduce dimensions with TICA - find the slow collective coordinates.
Cluster the TICA space into ~1000 microstates.
Build a Markov model: choose a lag time from the implied-timescales plot, then lump microstates into macrostates with PCCA.
Read kinetics: equilibrium populations, free-energy surface, mean first-passage times.
Setup#
import os
from glob import glob
from pathlib import Path
from htmd.ui import (
simlist, simmerge,
Metric, MetricSelfDistance,
TICA, Model, Kinetics,
)
from sklearn.cluster import MiniBatchKMeans
Please cite HTMD: Doerr et al.(2016)JCTC,12,1845. https://dx.doi.org/10.1021/acs.jctc.6b00049
HTMD Documentation at: https://software.acellera.com/htmd/
You are on the latest HTMD version (2.8.5.dev17+gf26491d44.d20260602).
2026-06-02 18:43:57,627 - rdkit - INFO - Enabling RDKit 2026.03.2 jupyter extensions
Step 1 - Build a simulation list#
The filtered trajectories (already stripped of waters and ions, aligned for analysis) ship as a zip on Figshare (HTMD tutorial data, DOI 10.6084/m9.figshare.32541291) - download protein_folding_datasets.zip and extract it once. ~2.5 GB. The tutorial reads the absolute path from the HTMD_TUTORIAL_DATASETS environment variable so the same notebook works regardless of where you extracted to.
The dataset is organised as several “epochs” (independent adaptive-sampling rounds). simlist() builds one list per epoch — duplicate folder basenames within a single call would raise — and simmerge() stitches the per-epoch lists into one combined list, renumbering the simid indices to be sequential across the merged whole:
DATASETS = Path(os.environ["HTMD_TUTORIAL_DATASETS"]) / "protein_folding_datasets"
topology = str(DATASETS / "1" / "filtered") # any epoch works - all share the same topology
sims = []
for epoch in sorted(DATASETS.glob("*/")):
trajs = glob(os.path.join(epoch, "filtered", "*", ""))
sims = simmerge(sims, simlist(trajs, topology))
len(sims)
2137
A Sim is the unit of analysis: trajectory frames + the matching topology / structure file (any moleculekit-supported topology format works — PDB, PSF, prmtop, …).
Step 2 - Project geometry to Cα distances#
Folding is encoded in the network of Cα-Cα contacts: in the unfolded state the helices are apart, in the folded state they pack against each other. MetricSelfDistance computes the upper-triangular distance matrix between every Cα pair, frame by frame. metric="contacts" thresholds those distances into 0/1 contacts (1 if below the threshold, 0 otherwise - default threshold=8 Å). Contact features are cheaper and more robust than raw distances for MSMs.
metr = Metric(sims)
metr.set(MetricSelfDistance("protein and name CA", metric="contacts", periodic="chains"))
data = metr.project()
data.fstep = 0.1 # ns per frame - tells downstream code the time axis
htmd.projections.metric - INFO - Frame step 0.1ns was read from the trajectories. If it looks wrong, redefine it by manually setting the MetricData.fstep property.
data is a MetricData object: per-trajectory projected coordinates plus the metric metadata.
data.plotTrajSizes()
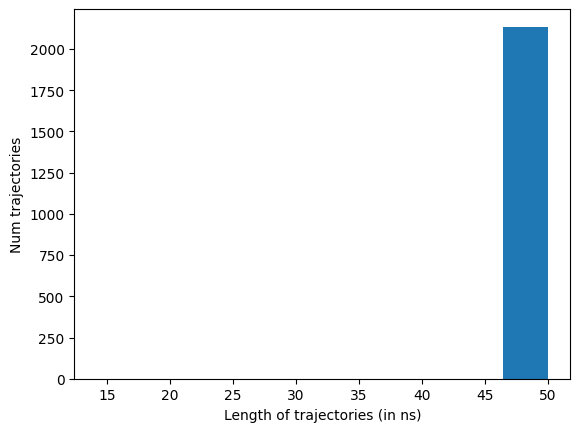
Drop any trajectories whose length isn’t the statistical mode (the most common frame count). By default dropTraj() removes anything off that mode — both crashed (too short) and over-run (too long) — so the rest of the pipeline sees a uniform-length set:
data.dropTraj()
htmd.metricdata - INFO - Dropped 7 trajectories from 2137 resulting in 2130
array([ 168, 472, 709, 1507, 1601, 2100, 2111])
Step 3 - TICA: find the slow coordinates#
TICA (Time-lagged Independent Component Analysis) re-projects the high-dimensional contact space onto the directions of slowest decorrelation. For an MSM, those slow directions are exactly what you want to discretise.
tica = TICA(data, 2, units="ns") # lag time for the TICA fit
dataTica = tica.project(3) # keep the 3 slowest TIC coordinates
The lag time you pick here (2 ns) should be a few times the fastest non-trivial relaxation; for villin folding 2 ns is plenty.
Step 4 - Cluster the TICA space#
Discretise the continuous TICA space into ~1000 microstates with mini-batch k-means. Bootstrap the data first (sample 80% of trajs) - re-running with different bootstrap draws gives you deviations / error bars on the timescales and is the standard check that the model is sample-stable:
dataBoot = dataTica.bootstrap(0.8)
dataBoot.cluster(MiniBatchKMeans(n_clusters=1000))
Step 5 - Build the MSM#
model = Model(dataBoot)
model.plotTimescales(maxlag=40, units="ns")
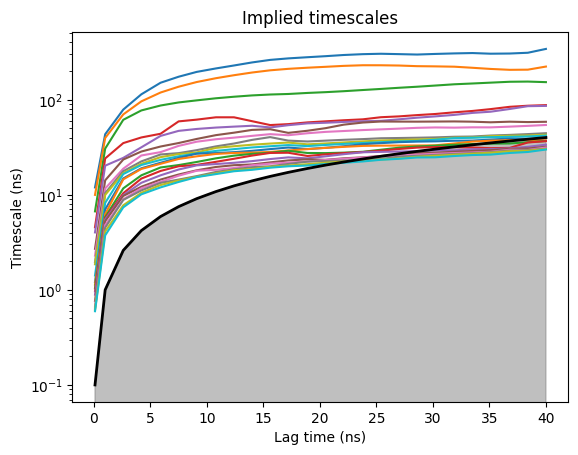
The implied-timescales (ITS) plot tells you when the model becomes Markovian: pick the shortest lag time at which the top timescales have plateaued. For villin folding that’s ~25 ns - the slow folding timescale is clearly separated from the bulk continuum.
model.markovModel(25, 4, units="ns")
htmd.model - INFO - 100.0% of the data was used
htmd.model - INFO - Number of trajectories that visited each macrostate:
htmd.model - INFO - [ 116 479 74 1655]
25 is the MSM lag time in ns; 4 is the number of macrostates PCCA lumps the microstates into. We pick 4 because the ITS plot shows the top 3 timescales clearly separated from the rest of the spectrum - 3 slow processes partition the microstate space into 3 + 1 = 4 metastable basins. The result captures the folded basin + a couple of unfolded sub-states + the transition region.
Step 6 - Equilibrium populations and FES#
model.eqDistribution()
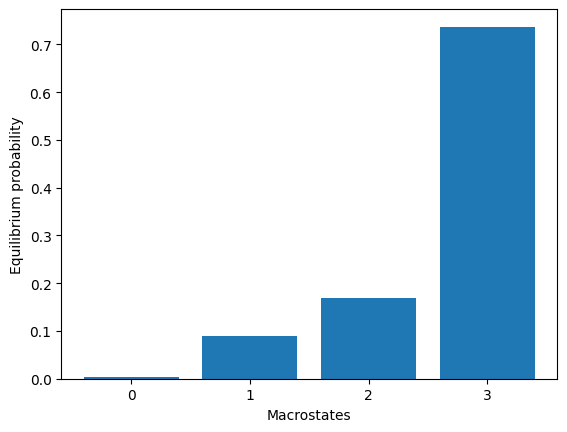
array([0.00353847, 0.09011645, 0.16917454, 0.73717054])
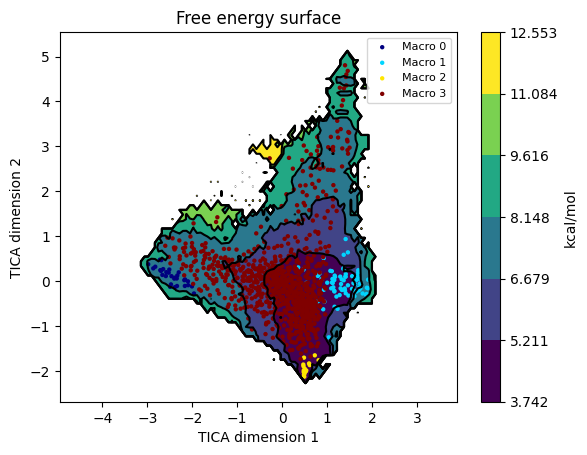
model.plotFES(0, 1, temperature=360)
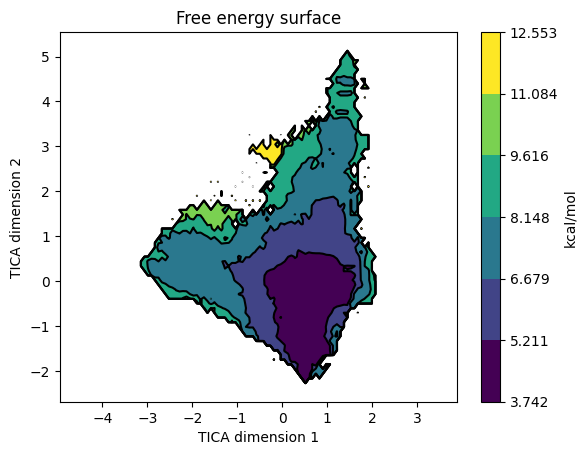
The FES on TIC1 / TIC2 shows the macrostate basins. Overlay the state assignments:
model.plotFES(0, 1, temperature=360, states=True)
To eyeball the macrostates as structures, run model.viewStates(protein=True) from an interactive session - it launches VMD with one representative frame per macrostate. (Omitted here because it needs a display.)
Step 7 - Kinetics#
kin = Kinetics(model, temperature=360)
r = kin.getRates()
print(r)
mfpton = 2.09E+03 (ns)
mfptoff = 2.35E+02 (ns)
kon = 4.79E+05 (1/M 1/s)
koff = 4.25E+06 (1/s)
koff/kon = 8.87E+00 (M)
kdeq = 4.36E+00 (M)
g0eq = 1.05 (kcal/M)
htmd.kinetics - INFO - Detecting source state...
htmd.kinetics - INFO - Guessing the source state as the state with minimum contacts.
htmd.kinetics - INFO - Source macro = 3
htmd.kinetics - INFO - Detecting sink state...
htmd.kinetics - INFO - Sink macro = 2
htmd.kinetics - INFO - Calculating rates between source: [np.int64(3)] and sink: [np.int64(2)] states.
Kinetics reads the macrostate model and computes mean-first-passage times and rates. getRates() returns a single Rates object for one auto-detected source → sink pair (folded → unfolded by default). To inspect all pairs, pass source= / sink= explicitly per call, or use kin.plotRates() (below) which iterates internally.
kin.plotRates()
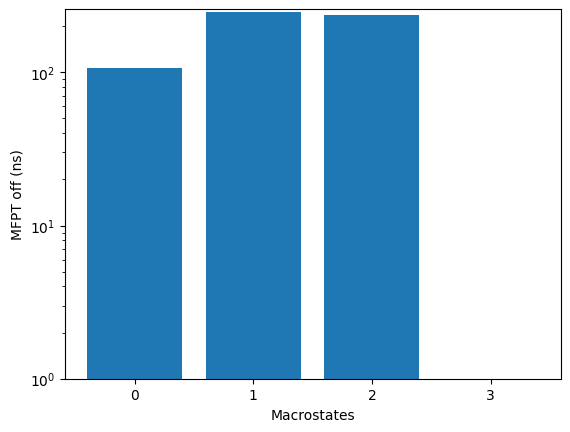
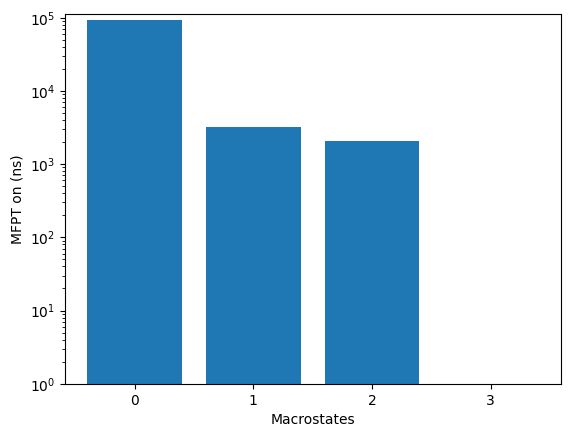
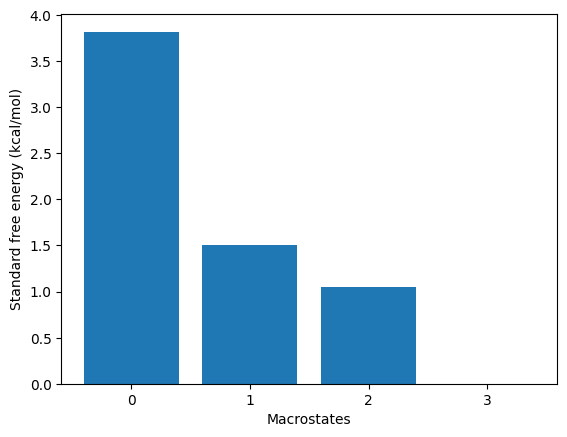
kin.plotFluxPathways()
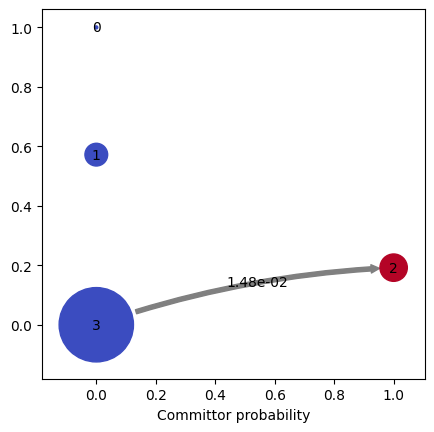
Path flux %path %of total path
0.014825165238110399 100.0% 100.0% [3 2]
Flux pathways visualise the dominant transition routes in TPT (transition-path theory) form - which intermediates the protein actually visits on its way from unfolded to folded.
Parameters that matter#
Knob |
Effect |
|---|---|
|
Contact features are coarser but more robust than raw distances - the binary thresholding makes clusters insensitive to small Cα-Cα fluctuations within a folded basin. |
|
Set a few times the fastest non-trivial relaxation. Too small → noise; too large → loses fast slow modes. |
|
Number of TIC coordinates kept. 3-5 is typical; check the eigenvalue spectrum to decide. |
|
More clusters = finer microstate resolution but slower. 500-2000 is the usual range. |
|
Read off the ITS plot - shortest lag at which slow timescales plateau. |
|
PCCA macrostates. Look at the timescale gaps in the ITS plot to pick the number. |
Gotchas#
ITS plateau, not minimum. Pick the lag where slow timescales first level off, not where they’re highest. Too-long lags throw away kinetic resolution for no gain.
bootstrap(0.8)lets you get deviations / error bars and see whether the model is sample-stable. Re-run with several bootstrap draws and compare; if different bootstraps give wildly different ITS / states, your dataset is undersampled.Macrostate count is a judgement call. Look at the timescale gap pattern - if you see a clear gap after k slow timescales, take k+1 macrostates.
See also#
MSM workflow explanation - the conceptual pipeline behind these calls.
Ligand binding MSM - the same pipeline for a binding system (uses
MetricDistancebetween protein and ligand instead ofMetricSelfDistanceon the protein).Adaptive sampling - how to generate the trajectory set that feeds this analysis.